|
GA's are widespread
and so far ubiquitous in both flowering (angiosperms) and non-flowering
(gymnosperms) plants as well as ferns.
Gibberellin
History
Gibberellin Biosynthesis and
Metabolism
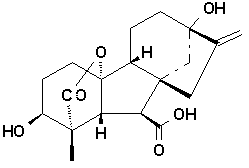
Gibberellins are diterpenes synthesized from
acetyl CoA via the mevalonic acid pathway. They all have either
19 or 20 carbon units grouped into either four or five ring systems.
The fifth ring is a lactone ring as shown in the structures above
attached to ring A. Gibberellins are believed to be synthesized
in young tissues of the shoot and also the developing seed. It is
uncertain whether young root tissues also produce gibberellins.
There is also some evidence that leaves may be the source of some
biosynthesis (Sponsel, 1995; Salisbury and Ross). The pathway by
which gibberellins are formed is outlined below and illustrated
in figure1.
3 acetyl CoA molecules are oxidized by 2 NADPH molecules to produce
3 CoA molecules as a side product and mevalonic acid.
Mevalonic acid is then Phosphorylated by ATP and decarboxylated
to form isopentyl pyrophosphate.
4 of these molecules form geranylgeranyl pyrophosphate which serves
as the donor for all GA carbon atoms.
This compound is then converted to copalylpyrophosphate which has
2 ring systems
Copalylpyrophosphate is then converted to kaurene which has 4 ring
systems
Subsequent oxidations reveal kaurenol (alcohol form), kaurenal (aldehyde
form), and kaurenoic acid respectively.
Kaurenoic acid is converted to the aldehyde form of GA12 by decarboxylation.
GA12 is the 1st true gibberellane ring system with 20 carbons.
From the aldehyde form of GA12 arise both 20 and 19 carbon gibberellins
but there are many mechanisms by which these other compounds arise.
Certain commercial chemicals which are used to stunt growth do so
in part because they block the synthesis of gibberellins. Some of
these chemicals are Phosphon D, Amo-1618, Cycocel (CCC), ancymidol,
and paclobutrazol. During active growth, the plant will metabolize
most gibberellins by hydroxylation to inactive conjugates quickly
with the exception of GA3. GA3 is degraded much slower which helps
to explain why the symptoms initially associated with the hormone
in the disease bakanae are present. Inactive conjugates might be
stored or translocated via the phloem and xylem before their release
(activation) at the proper time and in the proper tissue (Arteca,
1996; Sponsel, 1995).
Functions of Gibberellins
Active gibberellins show many physiological
effects, each depending on the type of gibberellin present as well
as the species of plant. Some of the physiological processes stimulated
by gibberellins are outlined below (Davies, 1995; Mauseth, 1991;
Raven, 1992; Salisbury and Ross, 1992).
- Stimulate stem elongation
by stimulating cell division and elongation.
- Stimulates bolting/flowering
in response to long days.
- Breaks seed dormancy
in some plants which require stratification or light to induce
germination.
- Stimulates enzyme
production (a-amylase) in germinating cereal grains for mobilization
of seed reserves.
- Induces maleness in
dioecious flowers (sex expression).
- Can cause parthenocarpic
(seedless) fruit development.
- Can delay senescence
in leaves and citrus fruits.
|